P2X受容体(P2Xじゅようたい、英: P2X receptors)は、陽イオン透過性イオンチャネル内蔵型受容体であり、細胞外アデノシン三リン酸(ATP)と結合して反応する受容体である。プリン受容体ファミリーに属する。P2X受容体はヒトのほか、マウス、ラット、ウサギ、ニワトリ、ゼブラフィッシュ、ウシガエル、吸虫、アメーバ等、多種多様な種に存在する。
生理学的役割
P2X受容体は下記の様な多彩な生理学的機能に関与している。
- 心臓のリズムと収縮性の調節
- 血管抵抗の調節
- 疼痛の伝達(特に慢性疼痛)
- 射精時の精管の収縮
- 排尿時の膀胱の収縮
- 血小板の凝集
- マクロファージの活性化
- アポトーシス
- 神経細胞とグリア細胞の相互作用
組織内の分布
P2X受容体は広く様々な動物種の組織に発現している。神経末端のシナプス前/後、ならびに中枢神経系、末梢神経系、自律神経系の全体にわたるグリア細胞では、P2X受容体はシナプス伝導を制御している。加えて、心筋や骨格筋のほか、血管、精管、膀胱等の平滑筋で、筋収縮の開始を司っている。P2X受容体はリンパ球やマクロファージ等の白血球あるいは血小板でも発現している。一部の細胞では発現しているP2X受容体のサブタイプに偏りが見られる。P2X1受容体は平滑筋細胞に目立ち、P2X2受容体は自律神経系に広く分布している。しかし、これらは極端に偏在しているものではなく、各サブユニットの分布は重なっており、ほとんどの細胞で2つ以上のサブユニットが発現している。例えば、P2X2とP2X3は感覚神経に共に発現しており、機能的には1つのものとしてP2X2/3受容体と呼ばれる。
基本構造と命名法
P2Xをコードする遺伝子には7種類が知られており、P2X1からP2X7と呼ばれている。
サブユニットのトポロジーは全て共通しており、細胞膜を貫通する部位を2つ有し、大きな細胞外ループで繋がっており、C末端とN末端は細胞内にある(図1)。N末端側にはプロテインキナーゼCのコンセンサス部位(キナーゼでリン酸化される部位)を含んでおり、P2Xサブユニットのリン酸化が受容体の機能の本質である可能性を示唆している。加えて、C末端側は変化に富んでおり、それぞれがサブユニット固有の機能の元となっている可能性が示される。
一般的に言うと、多くのサブユニットはホモマーまたはヘテロマーとして多量体を形成し、受容体となっており、受容体は構成されるサブユニットに基づいて命名される。すなわち、P2X1のみで構成されるホモマー受容体はP2X1受容体と、P2X2とP2X3から成るヘテロマー受容体はP2X2/3と呼ばれる。一般に、P2X6サブユニットはモノマー受容体を構成しても機能せず、P2X7サブユニットはヘテロマーでは機能しないと考えられている。
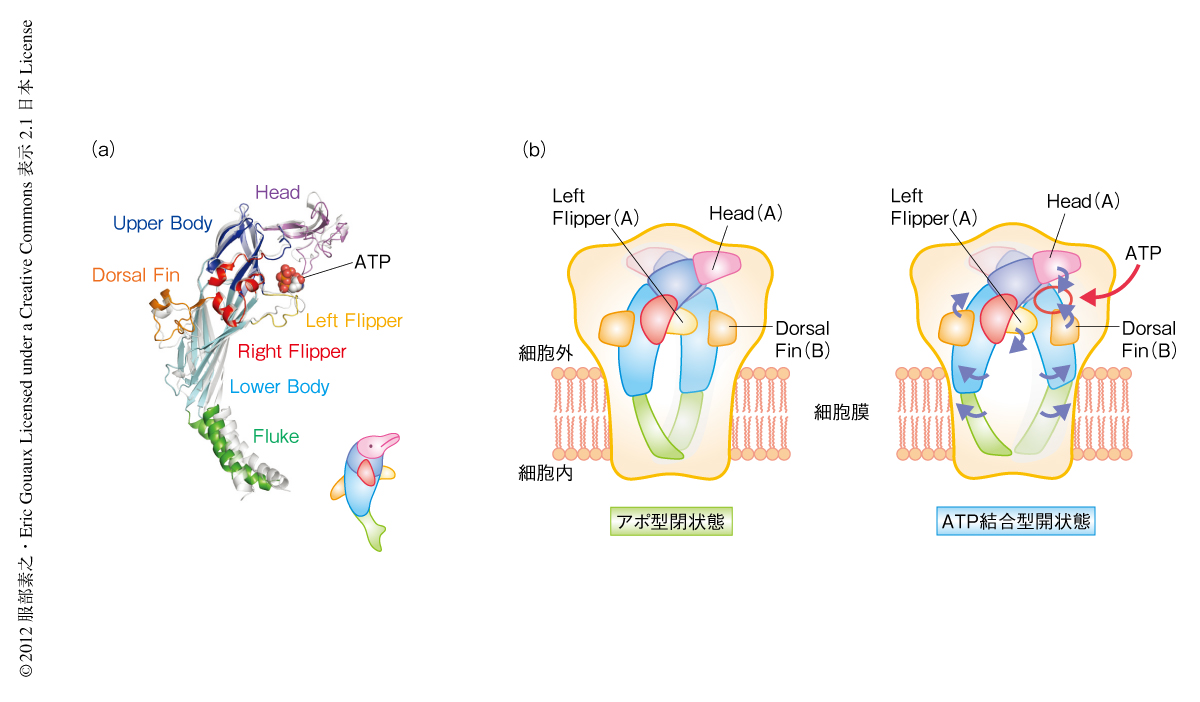
初期の分子生物学やタンパク質の機能的研究では、3本のペプチドサブユニットが会合して中央にイオン透過チャネル孔が形成されたので、機能を持つP2X受容体タンパク質が三量体であることを示す強い根拠であると考えられた。このことはゼブラフィッシュのP2X4受容体の三次元構造をX線結晶構造解析した結果からも裏付けられた(図2)。これらの知見は、各サブユニットの第2膜貫通ドメインがイオン透過孔を形作り、チャネルの開閉を司っていることを示している。
P2X受容体の構造と活性の相関は多くの研究の主題となり、その結果、ATP結合、イオン透過性、孔拡大、感受性低下を担う部位が特定された。
活性化とチャネル開口
P2Xの活性化には3分子のATPが必要であると考えられ、チャネル孔の開口には3つのサブユニットのそれぞれにATPが結合する必要があると思われていたが、その後、ATPは3つのサブユニットの境界部に結合することが判明した。ATPがP2X受容体の細胞外ループに結合すると、イオンチャネルのコンフォメーションの変化を引き起こし、イオン透過口を開口させる。最も受け入れられているチャネル開口のメカニズムは、第2膜貫通ドメイン(TM2)螺旋構造の回転と解離であり、3つのTM2が広がったその側面(孔)をNa やCa2 といった陽イオンが通過すると思われる。陽イオンが進入すると細胞膜の脱分極が起こり、数多くのCa2 応答性細胞内反応が惹起される。チャネルが開口している時間は受容体を構成するサブタイプに依存する。例を挙げると、P2X1受容体とP2X3受容体はATP存在下で急速(数百ミリ秒)に応答性が低下し、一方でP2X2受容体はATPが結合している限りチャネルが開いている。
薬理学上の役割
P2X受容体の薬理学的挙動は、主に受容体を構成するサブユニットの構成により定まる。異なるサブユニットは、ATP、α,β-メチレンATP、2'(または3')-O-(4-ベンゾイルベンゾイル)ATP(BzATP)等のプリン作動薬や、ピリドキサールリン酸-6-アゾフェニル-2',4'-ジスルホン酸(PPADS)やスラミン等のプリン拮抗薬に対して異なる感受性を有する。一部のP2X受容体(P2X2、P2X4、ヒトP2X5、P2X7)がATP応答性に多段階のチャネル開口反応を示し、時間経過と共に大きな有機陽イオン〔N-メチル-D-グルカミン(NMDG )〕やヌクレオチド結合性色素〔ヨウ化プロピジウム(YO-PRO-1)〕を透過する様になるという事実は引き続き興味の対象となっている。この透過性の変化がP2X受容体の孔径が大きくなる事によるものか他のイオン透過孔が開口する事によるものかとのテーマが研究途上にある。
生合成と輸送
P2X受容体は粗面小胞体で合成される。ゴルジ体で複雑な糖鎖が結合された後、受容体は細胞膜に運ばれてSNAREタンパク質ファミリーの一部と結合する。C末端のYXXXKという配列モチーフは全てのP2Xサブユニットに共通であり、サブユニットの輸送および細胞膜中でのP2X受容体の安定化にとって重要であると思われる。P2X受容体が細胞膜から除去される際には、クラスリン誘導性食作用によるエンドソーム(小胞の一種)へ取り込まれて、リソソームで分解、再利用される。
アロステリック調節
P2X受容体のATPに対する感度は細胞外のpHや重金属(亜鉛やカルシウム)の存在に強い影響を受ける。例えば、P2X1受容体、P2X3受容体、P2X4受容体のATP感受性は細胞外pHが7未満では弱く、P2X2受容体の感受性は著しく亢進する。その一方、亜鉛イオンはP2X2、P2X3、P2X4のATP誘導性イオン流を増強し、P2X1については阻害する。pHや重金属イオンのP2X受容体へのアロステリック効果は、細胞外ドメインのヒスチジン側鎖が授与している。P2X4受容体は他のP2X受容体とは対照的に、大環状ラクトンであるイベルメクチンに大きく影響される。イベルメクチンはATP存在時のP2X4チャネル開口の確率を上昇させて、ATP誘導性イオン流を増強し、脂質二重膜の内側から膜貫通ドメインに相互作用している様に見える。
サブファミリー
- P2RX1
- P2RX2
- P2RX3
- P2RX4
- P2RX5
- P2RX6
- P2RX7
P2Xドメインを含むヒトタンパク質
P2RX1、P2RX2、P2RX3、P2RX4、P2RX5、P2RX7、P2RXL1、TAX1BP3
薬理学
アゴニスト
- P2X4受容体増強
イベルメクチン、亜鉛
アンタゴニスト
- P2X4受容体阻害
パロキセチン(ラットP2X4受容体IC50=2.45µM、ヒトP2X4受容体IC50=1.87µM)、フルオキセチン、クロミプラミン、マプロチリン、その他抗うつ薬
一般的に抗うつ薬の典型的な血清濃度は、約100〜1,000nMの範囲。抗うつ薬は親油性の傾向があり組織に蓄積するので、中枢神経系では実験で観察されたP2X4受容体阻害に有効な濃度に到達する。
出典
関連項目
- イオンチャネル内蔵型受容体
外部リンク
- Ivar von Kügelgen: Pharmacology of mammalian P2X- and P2Y-receptors, BIOTREND Reviews No. 03, September 2008,© 2008 BIOTREND Chemicals AG
- Ligand-gated ion channel Database (European Bioinformatics Institute)